Vztah mezi strukturou rostlinného orgánu a jeho funkcí
Většina fyziologických procesů v rostlinách je přísně lokalizována:
fotosyntéza probíhá v chloroplastech,
transpirace má úzkou vazbu na rozložení, plošnou hustotu a otevřenost průduchů,
transportní procesy na dlouhé vzdálenosti se neobejdou bez specializovaných cévních
svazků...V rámci tohoto cvičení budete mikroskopovat dva rostlinné orgány, kořeny
a listy, jejichž
vnější a vnitřní uspořádání má úzký vztah k jejich fyziologickým
funkcím. Konkrétně budete pozorovat :
- endodermis, popřípadě hypodermis či exodermis kořene
- kořenové vlásky; úloha je spojená s výpočtem sorpční plochy kořenového
vlášení
- organizaci cévních svazků listů C4 rostlin
1. Lignifikace endodermis kořene, exodermis a hypodermis
Princip:
Endodermis se vyvíji v kořenech již v poměrně malé vzdálenosti za apikálním
meristémem. Může prodělat celkem tři vývojové fáze, ovšem už první z nich plně
zabezpečuje nejdůležitější funkci endodermis: tvoří bariéru volnému
apoplastickému pohybu vodných roztoků směrem do středního válce, což těsně
souvisí s příjmem živin.
Tato první vývojová fáze je charakteristická impregnací radiálních primárních
buněčných
stěn endodermis ligniny a suberiny, tedy látkami hydrofobní
povahy, měnícími buněčnou stěnu původně pro vodní roztoky propustnou
na
nepropustnou. Místa depozice suberinů a ligninů jsou na příčném řezu kořenem
viditelná jako tmavší místa, která nazýváme Casparyho proužky.
U některých rostlinných druhů
mohou být subepidermální vrstvy primární kůry anatomicky
odlišné od zbylých vrstev primární kůry (kortexu). Nazýváme je hypodermis, popř. exodermis. Termín
exodermis používáme, pokud buňky zevního kortexu procházejí
stejnými vývojovými fázemi jako endodermis. Pokud tomu tak není, používáme
termín hypodermis. Exodermis je vždy jednovrstevná, zatímco hypodermis bývá
vícevrstevná a může jako jednu z vrstev obsahovat exodermis; v tomto případě
mluvíme o složené hypodermis. Tak jako endodermis, i exodermis tvoří bariéru
volnému apoplastickému toku látek. Naproti tomu hypodermis má funkci
mechanickou a ochrannou.
Lignin v buněčných stěnách endodermis, exodermis či hypodermis budete dokazovat pomocí často
používaného histochemického barvení floroglucinolem (1% roztok floroglucinolu v 96%
etanolu). Reakce probíhá v kyselém prostředí, lignifikované buněčné stěny se
barví skořicově až sytě červeně.
Rostlinný materiál:
Endodermis: předklíčené, nejméně několik dní staré rostliny Zea mays, Vicia faba;
vzdušné kořeny Monstera deliciosa.
Exodermis a hypodermis: předklíčené, nejméně několik dní staré rostliny Zea
mays pěstované ve vodní kultuře; vzdušné kořeny Oncidium sp.,
popř. jiných epifytických orchidejí.
Vlastní provedení:
Od každého rostlinného druhu zhotovíme několik dostatečně kvalitních příčných
ručních žiletkových řezů, které shromažďujeme ve vodě na hodinovém sklíčku.
Poté odsajeme přebytek vody, přidáme několik kapek roztoku floroglucinolu a malou
kapku koncentrované HCl (okyselení roztoku). Necháme několik minut vybarvovat.
Preparáty montujeme do vody a bezprostředně pozorujeme - intenzita zabarvení se ve vodě
postupně snižuje. Preparáty ovšem nesmíme montovat do vlastního barvícího roztoku
- mohlo by dojít k naleptání objektivů mikroskopu kyselinou!
2. Pozorování kořenového vlášení, výpočet plochy sorpčního
povrchu kořene
Princip:
Kořenové vlášení (soubor kořenových vlásků, což jsou vychlípeniny buněk kořenové
epidermis) má důležitou roli v příjmu vody a minerálních živin z půdy. Podstatným způsobem
totiž zvyšuje objem půdy, z nichž jsou tyto živiny sorbovány. To má zásadní
význam
zejména u živin v půdě málo mobilních, jako jsou fosfátové ionty. Kořenové
vlášení navíc zvětšuje zevní povrch kořene, což opět vede ke zvýšené
sorpci vody a živin rostlinou. Délka, hustota a životnost
kořenových vlásků má tedy pro růst
rostlin zásadní význam. Kořenovým vlášením přitom není pokryt celý povrch všech
kořenů rostliny - kořenové vlásky vyrůstají a posléze odumírají v určité vzdálenosti od
apexu kořene. Rostliny rostoucí v zamokřených půdách (případně
pěstované hydroponicky) mívají redukované kořenové vlášení, které může
případně i zcela chybět.
Rostlinný materiál:
Předklíčené, několik dní staré rostliny Zea mays, Vicia faba, Pisum sativum,
Helianthus annuus...
Vlastní provedení:
Kořenové vlášení pozorujeme jednak v intaktním stavu pomocí binokuláru
(preparačního mikroskopu), jednak pomocí mikroskopické techniky světlého pole, kdy
pozorujeme jednotlivé příčné ruční řezy kořenů s kořenovým vlášením
namontované do
vody.
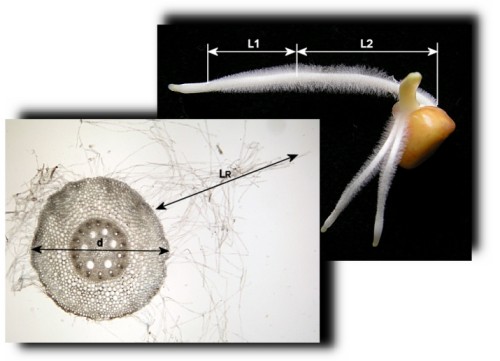 |
Pro výpočet plochy sorpčního povrchu kořene musíme zjistit následující
znaky kořene:
- délku zóny kořenového vlášení, kterou stanovíme s pomocí
milimetrového měřítka, v případě potřeby s využitím binokuláru, s
přesností na mm; stanovíme zvlášť délku zóny kořenového vlášení
s vyvíjejícími se vlásky (L1) a s plně vyvinutými vlásky (L2)
- průměr kořene ve středu zóny kořenového vlášení (d) a délku typického
plně vyvinutého kořenového vlásku (LR); zjistíme po zhotovení příčných
ručních řezů kořenem pomocí mikroskopu; hodnoty obou znaků odhadneme
z proporcí k průměru zorného pole mikroskopu při daném použitém
objektivu (hodnotu sdělí vyučující; d a LR odhadněte s přesností na desetiny
mm)
|
Pro výpočet budeme:
- uvažovat plošnou hustotu 20 kořenových vlásků
na 1 mm2 kořenové epidermis (bez povrchu kořenových
vlásků)
- závislost prodlužování kořenových vlásků v zóně L1 považovat
za lineární
- délku kořenových vlásků v zóně L2 považovat za konstantní
- kořenové vlásky považovat za válce o průměru 50 µm
K výpočtu využijte následující rovnice:
| Plocha rhizodermis (bez plochy kořenových vlásků) v zóně vyvíjejícího
se kořenového vlášení |
S1 = L1 × π × d [mm2] |
| Plocha rhizodermis (bez plochy kořenových vlásků) v zóně
plně vyvinutého kořenového vlášení |
S2 = L2 × π × d [mm2] |
| Celková plocha rhizodermis (bez plochy kořenových vlásků) |
SRH = S1 + S2 [mm2] |
|
Plocha kořenových vlásků v zóně kořenového vlášení
s vyvíjejícími se vlásky (zóna L1) |
SKV1 = S1 × 20 × (LR × π ×
0,05] × 0,5 [mm2] |
|
Plocha kořenových vlásků v zóně kořenového vlášení
s plně vyvinutými kořenovými vlásky (zóna L2) |
SKV2 = S2 × 20 × (LR × π
× 0,05] [mm2] |
|
Celková plocha kořenového vlášení |
SKV = SKV1 + SKV2 [mm2] |
|
Poměrné zvětšení zevního povrchu kořene o povrch kořenového vlášení |
SREL = (SKV / SRH) × 100
[%] |
3. Organizace cévních svazků C4 rostlin
Princip:
Většina rostlin na Zemi má tzv. C3 cyklus fotosyntetické asimilace CO2.
U některých rostlin, zejména v teplejších klimatických pásmech, se vyvinul
přídatný enzymatický aparát, který umožňuje tzv. Hatch-Slackovu dráhu fixace CO2,
kterou označujeme také jako C4. Výhoda tohoto fotosyntetického cyklu
spočívá v potlačení oxygenázové aktivity enzymu známého pod akronymem Rubisco,
tedy v potlačení fotorespirace. Jeho principem je prostorové oddělení primární
fixace CO2 z atmosféry v buňkách listového mezofylu (enzymatický aparát,
známý pod akronymem Pepco, má vysokou afinitu vůči CO2) od dekarboxylace a
následné finální fixace CO2 v buňkách pochev cévních svazků (nízká
afinita Rubisco vůči CO2, ovšem vysoký parciální tlak CO2).
Detailní chemismus C4 fixace CO2, stejně tak jako CAM
cyklus, je
probrán v rámci přednášek kurzu Fyziologie rostlin, popřípadě zkuste
tento anglický
text.
Rostlinný materiál:
Listy C4 rostlin: např. Zea mays, Cyperus papyrus, Saccharum
officinarium...
Vlastní provedení:
Od každého rostlinného druhu zhotovíme několik dostatečně kvalitních příčných
ručních žiletkových řezů, které shromažďujeme ve vodě na hodinovém sklíčku.
Namontujeme do vody a pozorujeme technikou světlého pole. Pozor! Cévní svazky s
pochvami buněk nemusí být úplně vyvinuté!
Prezentace výsledků
- Všechny pozorované preparáty zakreslete na listy bílého A4 papíru, jeden nákres
minimálně na polovinu A4 papíru. K nákresu použijte pouze tužku, kreslete pouze
obrysy struktur, nešrafujte ani nevykreslujte plochy. Popište všechny pozorované
struktury.
- V případě výpočtu sorpčních povrchů kořene, vyjádřete, o kolik
procent je zvětšen povrch kořene přítomností kořenových vlásků
oproti stavu, kdy kořenové vlásky chybí (SREL).