Primární anatomická stavba kořene
Teoretický souhrn
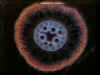
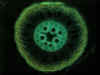
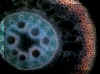
Povrch kořene je v typickém případě kryt jednovrstevnou rhizodermis (syn. kořenovou epidermis),
která může vytvářet kořenové vlásky. Vzdušné kořeny
epifytických rostlin obvykle mají pokožku vícevrstevnou, tzv. velamen (např.
epifytické orchideje).
Pod krycím pletivem se nachází zpravidla mnohovrstevná primární kůra
neboli kortex. Z hlediska příjmu vody a minerálních živin rostlinou má
největší význam nejvnitřnější vrstva primární kůry přiléhající ke střednímu
válci, označovaná jako endodermis. V závislosti na ontogenetickém
vývinu a taxonomické pozici rostliny mohou endodermální buňky prodělávat postupný vývoj a jejich
buněčné stěny mohou být postupně impregnovány suberinem, popřípadě mohou být i
lignifikovány. Pokud jsou v ranných fázích vývoje impregnovány pouze radiální
buněčné stěny endodermis, označujeme místa depozice suberinu a ligninu na příčném
řezu kořenem jako tzv. Casparyho proužky. V dalším vývoji jsou
často dále impregnovány a tloustnou vnitřní periklinální a radiální buněčné
stěny endodermálních buněk nebo tyto tloustnou a jsou impregnovány po celém
svém povrchu. Toto "utěsnění" jinak pro vodní roztoky volně
propustných buněčných stěn a intercelulár způsobí, že veškeré látky
přijímané rostlinou z půdního roztoku musí být před vstupem
do středního válce transportovány přes cytoplazmatickou membránu dovnitř buněk.
Tento přenos vyžaduje energii (obvykle se děje proti koncentračnímu gradientu dané
látky) a pro většinu látek se děje prostřednictvím specifických proteinových
přenašečů.
Zbylé buňky primární kůry jsou u starších kořenů, které ztrácejí
kořenové vlášení a jejichž sorpční funkce je potlačena, mohou být rozlišeny na
exodermis, tj. vrstvu analogickou endodermism avšak nacházející se bezprostředně pod rhizodermis, popř. na typicky vícevrstevnou hypodermis, tvořenou sklerifikovanými buňkami.
V centrální části kořene se nachází střední válec. S
endodermis sousedí jedno- nebo vícevrstevný pericykl, latentní
meristém, zakládající svou meristematickou činností boční kořeny. Vodivá pletiva kořene se nachází pod pericyklem a mají radiální uspořádání. Pro dvouděložné a nahosemenné
rostliny je typický nižší počet pólů cévního svazku (oligarchní radiální
organizace vodivých pletiv), naopak pro kořeny rostlin jednoděložných
je typické polyarchní
uspořádání vodivých pletiv, s vysokými počty protoxylémových pólů. Způsob zakládání cévních svazků je exarchní -
protoxylém se zakládá činností prokambia při obvodu středního válce, metaxylém
centripetálně od něj (tedy směrem do středu kořene). Uprostřed kořene se
může nycházet dřeň, tvořená typicky parenchymatickými buňkami, které
však mohou lignifikovat. U některých rostlin naopak dřeň chybí a v centrální
části středního válce se nachází elementy metaxylému.
Praktická část
Zhotovíme příčné ruční žiletkové řezy kořeny bobu setého (Vicia faba), harchu setého (Pisum sativum),
kosatcem (Iris sp.), případně kukuřicí (Zea mays), ječmenem (Hordeum
vulgare), vzdušným kořenem monstery (Monstera deliciosa) nebo některé
epifytické orchideje (Dendrobium, Phalaenopsis, Cattleya apod.). Lignifikované
buněčné stěny můžeme zvýraznit pomocí barvení floroglucinolem, popřípadě je možné preparáty pozorovat
s využitím autofluorescence, které je výhodné zejména k odlišení
morfologicky odlišných vrstev primární kůry.